Début de l'article <= |
Les égylopes provençales (I) ;
|
Accueil du site Contact~JM |
L'histoire des blés (Triticum sp.)
Presque au même moment et sur tous les continents les humains ont domestiqué des plantes céréalières : le riz en Asie, le maïs en Amérique du sud, le sorgho, le mil et le riz en Afrique, l'orge et les blés au moyen-orient et un peu plus tard en Europe.
Les blés (genre Triticum) sont des plantes de la famille des poacées. Le fruit caractéristique est un caryopse (*) communément appelé "grain". Les blés actuels sont cultivés pour produire du pain (Triticum æstivum) ou des pâtes et de la semoule (Triticum durum) mais leur histoire débute bien avant l'apparition de notre espèce.
Les données ethnobotaniques, cytologiques et génomiques ont permis de reconstituer une partie de cette histoire.
Vers − 0,5 Ma, dans la région du croissant fertile se sont produites les premières hybridations interspécifiques entre Triticum urartu et une espèce inconnue du groupe Sitopsis proche de Ægylops speltoides (Fig1) .
L'hybridation interspécifique réunit les génomes haploïdes (B) et (A) et conduit à un génome (AB) à 14 chromosomes mais les différences chromosomiques empêchent une gamétogenèse normale.
Une duplication complète rétablit la fertilité avec un caryotype à 2n = 4x = 28 chromosomes.
Le bilan est donc une addition des génomes (B) et (A) dans un organisme fertile de génome (AABB).
Cet organisme fertile est l'amidonier sauvage Triticum turgidum ssp dicoccoides.

▲ Fig1: Histoire des blés, modifié d'après (1) et (5).
(Dates indicatives variables selon les sources. 1 ka = 1000 ans)
Une deuxième hybridation naturelle avec une égylope diploïde (2n = 14) suivie d'une duplication conduit à la formation d'un organisme hexaploïde (2n = 6x = 42) de génome (AABBDD).
Cette dernière hybridation naturelle (T. turgidum x Æ. tauschii ) a dû se faire avec un petit nombre de géniteurs sauvages si bien que la diversité du génome (D) dans les blés hexaploïdes actuels est réduite.
Or les égylopes actuelles (Fig2) confrontées naturellement aux pathogènes et à des climats variés peuvent constituer une ressource génétique utile pour les blés cultivés.

▲ Fig2: Morphologie des épis des égylopes à génome (D) d'après (2).
(Æ. tauschii ssp. strangulata (a), Æ. tauschii ssp. tauschii (b), Æ. crassa (c),
Æ. cylindrica (d), Æ. juvenalis (e), Æ. vavilovii (f), Æ. ventricosa (g))
Ainsi chez le blé tendre, la variété lignée pure RENAN, obtenue en 1980 par l'INRA présente 5 gènes de résistance aux pathogènes. Ces résistances limitent les traitements phytosanitaires et rendent la variété exploitable en agriculture biologique. Les gènes de résistance viennent du génome (D) de Æ. ventricosa.
En effet dans la généalogie de RENAN le croisement qui apporte des gènes de résistance est le croisement artificiel Æ. ventricosa x T. persicum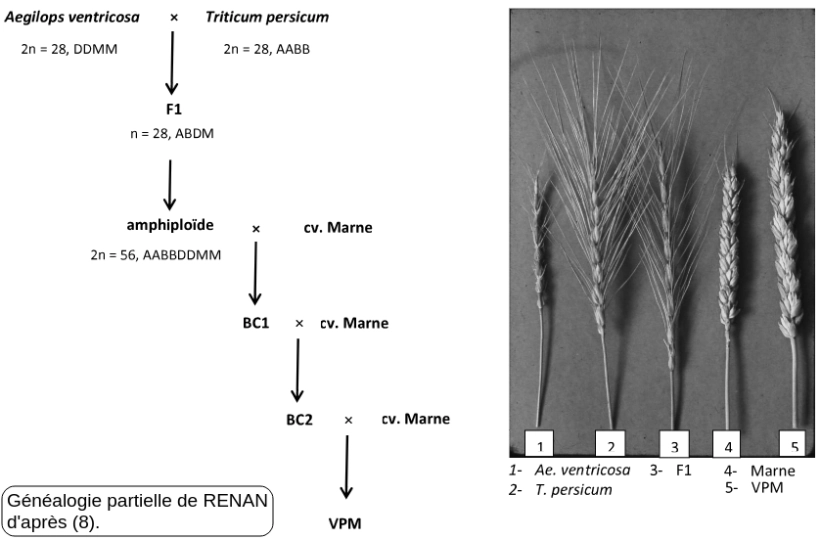
Parmi les égylopes à génome (D), Æ. ventricosa est un organisme polyploïde à génome additif de type (DDNN) issu de l'hybridation interspécifique Æ. tauschii x Æ. uniaristata (3).
C'est donc une espèce allotétraploïde (2n = 4x = 28) apparentée par Æ. tauschii aux blés actuels.

Relation de parenté entre les blés et l'égylope ventrue.
L'addition des génomes et la sélection artificielle, empirique puis scientifique ont considérablement modifié le phénotype des épis (Fig3).

▲ Fig3: Comparaison des épis d'égylopes et de blé.
(g = Æ. geniculata, v = Æ. ventricosa, b = T. æstivum)
La sélection a conduit au syndrome de domestication (**) : de nombreux grains par épi et de gros grains. Mais surtout un rachis solide qui ne se désarticule pas et donc ne disperse pas les grains et des grains nus auxquels les glumelles n'adhèrent pas. Ces deux derniers caractères sont contrôlés par l'allèle dominant du Gène Q.
Les égylopes (Ægylops sp.) provençales
On rencontre en Provence les deux égylopes Ægylops geniculata (ex ovata) et Æ. ventricosa qui forment souvent des peuplements monospécifiques (Fig4).
")
▲ Fig4: Stations où l'une des espèces domine (g = Æ. geniculata, v = Æ. ventricosa).
Néanmoins dans certaines stations les deux espèces peuvent se rencontrer (Fig5). Dans cette situation la proximité facilite les hybridations interspécifiques.

▲ Fig5: Station mixte des deux égylopes provençales (g = Æ. geniculata, v = Æ. ventricosa).
Parfois les peuplements de Æ. geniculata présentent une diversité phénotypique (Fig6). La forme typique est basse (20 cm) à épis vert-gris alors que l'autre forme est haute (30 cm) à épis vert-jaune.

▲ Fig6: Diversité phénotypique chez Ægylops geniculata.
(flèches = forme haute Æ. geniculata H et forme basse Æ. geniculata B)
L'origine de cette diversité nous est inconnue. Si on écarte un effet du milieu abiotique puisque les plantes poussent ensemble, il peut s'agir d'une interaction biotique (pathologie ou symbiose) ou bien d'une variation génétique (génotypique ou caryotypique).
Pour étudier les populations d'égylopes provençales nous avons recherché un indicateur du niveau de ploïdie plus facilement observable que le nombre de chromosomes.
Dimensions du grain de pollen et niveau de ploïdie
Les 2 égylopes provençales Ægylops geniculata (ex ovata) et Æ. ventricosa sont tétraploïdes (2n = 4x = 28) (4) alors que le blé cultivé est hexaploïde (2n = 6x = 42).
Chez l'avoine (Avena sp.) les dimensions du grain de pollen sont bien corrélées avec le niveau de ploïdie (6) qui varie de 2x à 8x comme chez les triticées.
Nos observations sur des grains de pollen prélevés à l'anthèse ont révélé une grande diversité dans les dimensions (Fig7).

▲ Fig7: Écart maximum observé dans les dimensions des grains de pollen.
(V = égylope ventrue (48 µm), B = blé tendre sans barbe (77 µm)
Flèches : blanche = pore, rouge = déformations symétriques, barre = 50 µm)
Les mesures indiquent des différences significatives dans la plus grande dimension (Fig8). Ces mesures sont assez bien corrélées avec les niveaux de ploïdie et la quantité d'ADN nucléaire en picogramme (pg) publiés (7).
▲ Fig8: Moyenne de la plus grande dimension (Dmax en µm) des grains de pollen (k = 69)
| Espèce | Dmax ± ICf5% (µm) | Ploïdie (2n) | QADN (pg) |
| Triticum æstivum | 66,5 ± 2,1 (a) | 42 | 17,7 (d) |
| Æ. geniculata H | 64,0 ± 0,9 (a) | - | - |
| Æ. geniculata B | 58,3 ± 1,3 (b) | 28 | 10,3 (e) |
| Ægylops ventricosa | 53,9 ± 1,2 (c) | 28 | 10,6 (e) |
ICf5% = intervalle de confiance t5%, ddl=68 (**)
Différences significatives : (a),(b),(c) pour Dmax, (d),(e) pour QADN.
En effet, nos mesures montrent que Dmax est significativement plus élevée pour Æ. geniculata H que pour
Æ. geniculata B et Æ. ventricosa . La valeur moyenne se rapproche de la valeur trouvée pour le blé tendre qui est hexaploïde. Ces résultats suggèrent donc que la forme haute puisse être une forme polyploïde ayant un nombre de chromosomes supérieur à 28.
Bilan:
La très ancienne histoire des blés est marquée par des évènements naturels d'hybridation interspécifique suivis de duplication complète. Ces 2 phénomènes conduisent à une addition des génomes parentaux. La sélection empirique, ancienne elle aussi, a porté sur des caractères morphologiques favorables à l'humain et entraîné le syndrome de domestication.
Plus récemment les Ægylops ont permis par des croisements interspécifiques d'introduire des gènes de résistance aux maladies dans les blés cultivés (Jahier & Rolland 2020, (8)).
On rencontre en Provence les deux espèces ; Ægylops geniculata (ex ovata) et Æ. ventricosa. Les populations de Æ. geniculata présentent une diversité morphologique et nos résultats suggèrent une différence caryotypique. Il peut donc s'agir pour la forme haute d'un hybride interspécifique spontané dont nous ne connaissons pas l'un des géniteurs.
Des hybrides naturels entre le blé tendre et Æ. geniculata ont été décrits (9). Néanmoins cela nous semble peu probable pour les plantes que nous avons observé car les épis sont très différents de ceux montrés par les auteurs.
Une caractérisation ultérieure, biométrique et caryotypique des égylopes que nous avons observé doit être réalisée.
Sur la domestication des céréales et plus généralement des plantes voir aussi le cours et le complément de cours.
Remarques:
▲ (*) Un caryopse est un fruit sec indéhiscent (qui ne s'ouvre pas) dont les parois sont soudées au tégument de la graine.
▲ (**) Syndrome de domestication : ensemble de caractères observés simultanément et qui sont à la fois favorables à l’humain et défavorables à la plante dans son milieu naturel.
▲ (***) Note technique sur le calcul des intervalles de confiance :
Les statistiques utilisées ici ont été calculées de manière semi-automatique avec Libre-Office-Calc sous Linux.
Nous avons calculé la moyenne [m = Somme / k, avec k = nombre de mesures] puis la Somme des Carrés des Écarts [SCE] et la variance empirique S2 = [SCE / (k-1)].
L'intervalle de confiance bilatéral au seuil de P/2 = 2,5%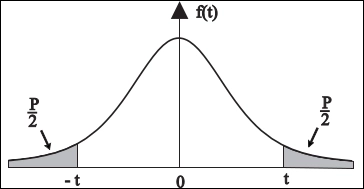 a été déterminé avec une table de la loi de Student au niveau (k-1) ddl et l'écart-type empirique S.
a été déterminé avec une table de la loi de Student au niveau (k-1) ddl et l'écart-type empirique S.
Remerciements :
▲ Nous tenons à remercier nos élèves de Spécialité_SVT_2023 qui ont courageusement et avec soin mesuré les dimensions de 69 grains de pollen lors de travaux pratiques.
Références: